오스트랄로피테쿠스 세디바
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
오스트랄로피테쿠스 세디바(Australopithecus sediba)는 2008년 남아프리카 공화국 말라파 동굴에서 발견된 화석을 바탕으로 2010년 명명된 오스트랄로피테쿠스 속의 종이다. 이 종은 골반과 다리에서 호모 에르가스터/H. 에렉투스와 유사한 특징을 보이며, 오스트랄로피테쿠스와 호모 사이의 전이 화석으로 여겨지기도 한다. A. 세디바는 뇌 용량이 작고, 긴 팔과 수목 생활에 적합한 특징을 지녔지만, 두 발 보행을 하였으며, C3 숲 식물을 섭취하고 뼈 종양과 외상을 입은 흔적이 발견되기도 했다. 말라파 유적지에서 발견된 동물 화석과 식물 규소체 분석을 통해 당시 환경을 유추할 수 있으며, A. 세디바는 파란트로푸스 로부스투스, 호모 에르가스터/H. 에렉투스와 공존했을 것으로 추정된다.
더 읽어볼만한 페이지
- 오스트랄로피테쿠스 - 타웅 아이
타웅 아이는 레이먼드 다트가 발견한 약 3~4세의 *오스트랄로피테쿠스 아프리카누스* 두개골 화석으로, 초기에는 논쟁이 있었으나 후속 연구를 통해 인류 진화 연구에 중요한 초기 호미닌으로 인정받았으며, 두개골 형태와 대후두공 위치는 직립 보행 가능성을 제시한다. - 오스트랄로피테쿠스 - 오스트랄로피테쿠스 가르히
약 250만 년 전 에티오피아에서 발견된 오스트랄로피테쿠스 가르히는 450cc 정도의 뇌 용적과 큰 어금니, 소구치를 가지며, 초기 도구 사용 흔적과 함께 발견되어 도구 사용과의 연관성이 제시되고, 종명은 아파르어로 "놀라움"을 뜻한다. - 아프리카의 고대 포유류 - 오스트랄로피테쿠스 가르히
약 250만 년 전 에티오피아에서 발견된 오스트랄로피테쿠스 가르히는 450cc 정도의 뇌 용적과 큰 어금니, 소구치를 가지며, 초기 도구 사용 흔적과 함께 발견되어 도구 사용과의 연관성이 제시되고, 종명은 아파르어로 "놀라움"을 뜻한다. - 아프리카의 고대 포유류 - 오스트랄로피테쿠스 아프리카누스
오스트랄로피테쿠스 아프리카누스는 약 300만 년 전에서 200만 년 전 사이 남아프리카에서 살았던 멸종된 사람족으로, 레이몬드 다트에 의해 발견되었으며 직립 보행을 했지만 유인원과 인간의 중간 형태를 보이는 인류 진화 연구에 중요한 종이다. - 2010년 기재된 포유류 - 월리스안경원숭이
월리스안경원숭이는 2010년 처음 기술된 술라웨시섬 고유종 안경원숭이로, 독특한 소리와 유전적 차이를 보이며 서식지 파괴로 인해 IUCN "취약" 등급으로 평가되었다. - 2010년 기재된 포유류 - 북부담황색뺨긴팔원숭이
북부담황색뺨긴팔원숭이는 수컷은 검은색 털과 밝은 갈색 가슴, 짙은 금빛-오렌지색 뺨을 가지며 암컷은 오렌지-베이지색 털을 가진 종이다.
| 오스트랄로피테쿠스 세디바 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 속 | 아우스트랄로피테쿠스속 |
| 종 | 아우스트랄로피테쿠스 세디바 |
| 명명자 | Berger 외, 2010 |
| 키 | 130cm |
| 키 (추정) | 150–156cm |
| 몸무게 | 30–36kg |
| 화석 정보 | |
| 시대 | 초기 홍적세, 1.98백만년 전 |
2. 연구사
오스트랄로피테쿠스 세디바의 연구는 2008년 8월 남아프리카 공화국 가우텡주의 인류의 요람 세계유산지역 내 말라파 동굴(Malapa Cave)에서 시작되었다. 고인류학자 리처드 로저스 버거(Lee Rogers Berger)의 당시 9세 아들인 매튜 버거(Matthew Berger)가 탐사 중 우연히 어린 개체의 쇄골 화석(MH1)을 발견한 것이 시초였다.[4] 이후 추가 발굴을 통해 MH1의 부분 골격과 또 다른 개체인 성체 암컷(MH2)의 화석이 발견되었다.[4]
발견된 화석들은 약 198만 년 전 초기 플라이스토세 시기의 것으로 연대가 측정되었다.[2][3] 2010년, 리 버거와 동료 연구자들은 이 화석들을 새로운 고인류 종으로 보고, MH1을 모식 표본(holotype)으로, MH2를 부모식 표본(paratype)으로 지정하여 ''오스트랄로피테쿠스 세디바''(''Australopithecus sediba'')라는 학명을 부여했다. 종명 "세디바"(''sediba'')는 남아프리카 공화국의 세소토어로 "샘" 또는 "우물"을 의미한다.[4]
''A. 세디바''는 발견 초기부터 오스트랄로피테쿠스와 후대의 호모 속의 특징을 함께 가지고 있어 주목받았다. 특히 골반과 다리 등에서 호모 속과 유사한 특징이 나타나, ''호모'' 속의 직계 조상이거나 가까운 친척일 가능성이 제기되며 인류 진화 연구에서 중요한 전이 화석 후보로 떠올랐다.[4][5] 그러나 화석의 연대가 이미 알려진 초기 ''호모'' 속 화석보다 후대라는 점 등 때문에 ''A. 세디바''의 정확한 계통학적 위치에 대해서는 현재까지도 학계에서 활발한 논의가 이루어지고 있다.[4][14][2]
2. 1. 표본
가장 먼저 발견된 표본은 MH1 (UW88-1)으로, 2008년 8월 15일 남아프리카 공화국 인류의 요람 내 말라파 동굴(Malapa Cave)에서 발견된 오른쪽 쇄골이다. 이는 고인류학자 리처드 로저스 버거(Lee Rogers Berger)의 9살 아들인 매튜 버거(Matthew Berger)가 발굴 현장을 탐사하던 중 우연히 발견했다.[4] 추가 발굴을 통해 MH1의 부분 골격이 더 발견되었는데, 여기에는 부분 두개골과 턱뼈 조각, 팔, 손가락, 어깨, 흉곽, 척주, 골반, 다리, 발 등이 포함되었다. MH1은 뚜렷하게 발달된 눈썹 융기, 송곳니, 치근, 턱뼈 각도의 외반, 뼈의 큰 흉터 등을 근거로 어린 수컷으로 해석되었다.[4] 그러나 인류학자 윌리엄 킴벨(William Kimbel)과 요엘 락(Yoel Rak)은 이러한 특징들이 성별을 결정하는 데 신뢰하기 어렵다고 주장하며, ''오스트랄로피테쿠스 아프리카누스(Australopithecus africanus)''의 성별 판별법을 적용하여 앞쪽 기둥(코 구멍에서 입 주위까지 내려오는 기둥)의 부재와 약간 볼록한 하비갑판을 근거로 MH1이 암컷일 수 있다고 제안했다.[14] MH1은 요하네스버그 세인트 메리 학교(St Mary's School)의 17세 학생 옴페메체 키필레(Omphemetse Keepile)에 의해 "카라보"(Karabo)라는 별명을 얻었다. 이는 츠와나어로 "답"을 의미하며, 키필레는 "이 화석이 인류 기원을 이해하는 데 대한 해답을 제시한다"는 의미로 이 이름을 선택했다.[1]
또 다른 주요 표본인 MH2는 성체 암컷으로 추정되며, 2008년 9월 4일 리처드 버거에 의해 발견되었다. MH2는 분리된 위쪽 치아, 부분 턱뼈, 거의 완전한 오른쪽 팔, 오른쪽 견갑골, 어깨, 척추 일부, 갈비뼈, 골반, 무릎 관절, 발 조각 등으로 구성된다. 넓고 사각형인 치골 형태와 약하거나 보통 수준의 근육 흉터는 MH2가 암컷임을 시사한다.[4]
| 표본 번호 | 별명 | 발견 날짜 | 발견자 | 추정 성별/나이 | 주요 발견 부위 |
|---|---|---|---|---|---|
| MH1 (UW88-1) | 카라보 (Karabo)[1] | 2008년 8월 15일 | 매튜 버거 | 어린 수컷 (논쟁 있음)[4][14] | 부분 두개골, 턱뼈 조각, 팔, 손가락, 어깨, 흉곽, 척주, 골반, 다리, 발[4] |
| MH2 | - | 2008년 9월 4일 | 리처드 로저스 버거 | 성체 암컷[4] | 고립된 위쪽 치아, 부분 턱뼈, 거의 완전한 오른쪽 팔, 오른쪽 견갑골, 어깨, 척추, 갈비뼈, 골반, 무릎 관절, 발 조각[4] |
표본이 발견된 지층은 초기 플라이스토세 시기에 형성된 것으로 추정된다. 이 지층을 덮고 있는 유석에 대한 우라늄-납 연대 측정 결과는 202.6만 ± 2.1만 년 전으로 나타났다.[3] 또한, 고지자기 연대 측정 결과 퇴적물이 현재와 같은 정상 자기장 방향(정상 잔류 자화)을 가지고 있어, 이 시기에 해당하는 195만 년 전에서 178만 년 전 사이에 퇴적되었음을 시사한다.[3] 2011년에는 유석을 이용한 우라늄-납 연대 측정을 다시 수행하여 197.7만 ± 2천 년 전이라는 더 정밀한 연대를 얻었다.[2]
2010년, 리 버거와 동료 연구자들은 MH1을 모식 표본(holotype)으로, MH2를 부모식 표본(paratype)으로 지정하여 ''오스트랄로피테쿠스 세디바''(''Australopithecus sediba'')라는 새로운 종을 공식적으로 발표했다. 종명 "''세디바''"는 현지 세소토어로 "샘" 또는 "우물"을 의미한다.[4]
2. 2. 분류
2010년, 리 버거(Lee Berger)와 동료 연구자들은 MH1 개체를 모식 표본(holotype)으로, MH2 개체를 부모식 표본(paratype)으로 지정하여 ''오스트랄로피테쿠스 세디바''(''Australopithecus sediba'')라는 새로운 종을 공식적으로 발표했다. 종명인 "세디바"(''sediba'')는 남아프리카 공화국의 세소토어(Sesotho language)로 "샘" 또는 "우물"을 뜻한다.[4] ''A. 세디바''는 특히 골반과 다리 부분에서 후대의 호모 에르가스터(Homo ergaster)나 호모 에렉투스(Homo erectus)와 유사한 특징을 많이 가지고 있어, 연구자들은 이 종이 오스트랄로피테쿠스(Australopithecus) 속과 호모(Homo) 속 사이의 전이 화석일 가능성이 있다고 보았다.[4] 치아의 특징 역시 ''A. 세디바''가 ''호모'' 속의 조상과 가까운 관계일 수 있음을 시사한다.[5]그러나 ''A. 세디바'' 화석이 발견된 지층은 약 195만 년 전에서 178만 년 전 사이의 것으로 추정되는데, 이는 당시 알려진 가장 오래된 ''호모'' 속 화석인 에티오피아 하다르(Hadar, Ethiopia) 지역의 ''호모 하빌리스(Homo habilis)''(약 233만 년 전)보다 훨씬 후대의 것이다.[4] 현재 가장 오래된 ''호모'' 속 표본은 에티오피아 레디-게라루(Ledi-Geraru) 지역에서 발견된 약 280만 년에서 275만 년 전의 LD 350-1 턱뼈 화석이다.[6] 이러한 연대 차이를 설명하기 위해, 연구자들은 ''A. 세디바''가 같은 지역에 살았던 더 오래된 종인 ''오스트랄로피테쿠스 아프리카누스(Australopithecus africanus)''의 한 개체군에서 진화했으며, 이후 ''호모'' 속이 ''A. 세디바''에서 갈라져 나왔을 것이라는 가설을 제시했다.[4] 이 가설은 ''A. 아프리카누스''와 ''A. 세디바'' 사이에 약 80만 년 동안 화석 기록이 발견되지 않는 유령 계통(ghost lineage)이 존재함을 가정한다.[14] 또한, 2011년에 말라파 동굴의 ''A. 세디바'' 화석 연대가 약 198만 년 전으로 더 정확히 측정되면서, 이는 당시 알려진 가장 오래된 ''H. 에르가스터''/''H. 에렉투스'' 화석보다 시기적으로 앞서는 것이었다. 이를 근거로 일부 연구자들은 ''H. 하빌리스''나 ''호모 루돌펜시스(Homo rudolfensis)'' 대신 ''A. 세디바''가 ''H. 에르가스터''/''H. 에렉투스''(논란의 여지가 적은 가장 오래된 ''호모'' 속 구성원)의 직접적인 조상일 수 있다고 주장했다.[2] 현재 ''A. 세디바''는 인류의 요람 지역에서 ''H. 에르가스터''/''H. 에렉투스'' 및 ''파란트로푸스 로부스투스(Paranthropus robustus)''와 같은 다른 고인류 종들과 동시대에 살았던 것으로 여겨진다.[7]
반면, ''A. 세디바''가 ''호모'' 속과는 직접적인 관련이 없고, ''A. 아프리카누스''의 후기 생존 형태이거나 단순히 자매 종일 가능성도 제기된다. 이 경우, ''A. 세디바''가 보이는 ''호모'' 속과 유사한 특징들은 수렴 진화의 결과, 즉 동형성(homoplasy)에 해당할 수 있다.[14][8][9][10][11] 초기 ''호모'' 속의 화석 기록은 매우 부족하고 단편적인 유물에 의존하는 경우가 많아, 해부학적 특징을 명확히 비교하기 어려운 점이 이러한 논쟁을 심화시킨다.[11] ''A. 아프리카누스'' 외에도 ''오스트랄로피테쿠스 아파렌시스(Australopithecus afarensis)''나 ''오스트랄로피테쿠스 가르히(Australopithecus garhi)'' 역시 ''호모'' 속의 조상 후보로 제안되어 왔으며, 어떤 종이 진정한 인류의 직계 조상인지는 여전히 학계의 주요 논쟁거리이다.[6] 또한, ''A. 세디바''의 모식 표본인 MH1이 어린 개체라는 점도 문제로 지적된다. 일부 학자들은 MH1에서 보이는 ''호모''와 유사한 얼굴 특징 중 일부는 성체로 성장하면서 사라졌을 수도 있다고 주장한다.[14]
현재 오스트랄로피테신(australopithecine)의 분류 체계는 매우 복잡하고 혼란스러운 상태이다. ''오스트랄로피테쿠스'' 속은 특정 공통 조상에서 유래한 단계통군이라기보다는, 유사한 생리적 특징을 공유하는 여러 종들을 편의상 묶어놓은 진화 등급(grade taxon)으로 간주되기도 한다. 따라서 대부분의 오스트랄로피테신 종들 사이의 정확한 계통 발생 관계는 아직 명확하게 밝혀지지 않았다.[12]
3. 해부학적 특징
오스트랄로피테쿠스 세디바는 고대 인류 조상인 오스트랄로피테쿠스 속의 특징과 더 진화된 호모 속의 특징을 함께 보여주는 독특한 해부학적 구조를 가진다. 이러한 여러 특징이 섞인 모습 때문에 인류 진화 과정에서 중요한 위치를 차지하는 종으로 여겨진다.
전체적으로 두 발 보행에 적응한 모습을 보이지만, 여전히 수목 생활에도 능숙했음을 시사하는 신체 구조를 유지하고 있었던 것으로 보인다. 예를 들어, 두개골은 뇌 용량이 작아 다른 오스트랄로피테쿠스와 유사하지만, 얼굴의 돌출 정도가 덜하고 치아가 작은 점 등은 초기 호모 속과 닮은 특징이다. 몸통의 경우, 허리뼈는 유연하여 직립 보행에 유리했지만, 상체는 유인원처럼 좁은 형태를 가졌다. 반면 골반은 초기 호모 속과 유사한 특징을 많이 공유한다.
팔은 상대적으로 길고 어깨 구조는 나무 타기에 적합한 유인원의 특징을 보여주지만, 손은 엄지손가락이 길고 나머지 손가락이 짧아 도구를 정교하게 다룰 수 있는 정밀 파지가 가능했을 것으로 추정된다. 다리는 전반적으로 직립 보행에 적응했지만, 발목과 발꿈치뼈 등 일부 구조에서는 유인원과 유사한 특징이 남아 있어, 완전히 현대 인류와 같은 방식의 보행은 아니었을 가능성을 시사한다.
이처럼 오스트랄로피테쿠스 세디바의 해부학적 특징은 과도기적 형태를 보여주며, 이는 인류가 어떻게 땅 위에서의 두 발 보행과 나무 위에서의 수목 생활 사이에서 적응해 나갔는지에 대한 중요한 단서를 제공한다.
3. 1. 두개골



MH1 개체는 두개골 천장만 보존되었으며, 그 부피는 363cm3이다. 뇌의 가장 뒷부분(7cm3~10cm3 추정)과 소뇌(다른 오스트랄로피테신 및 초기 호모 속과의 비교를 통해 약 50cm3로 추정)를 고려하면, MH1의 전체 뇌 부피는 약 420cm3~440cm3였을 것으로 추정된다. 이는 오스트랄로피테신에게서 일반적으로 나타나는 뇌 부피 범위에 속한다.[4] 현대 영장류의 성체와 신생아 뇌 크기 비율을 적용하면, ''A. 세디바'' 신생아의 뇌 크기는 153cm3~201cm3였을 것으로 추정되며, 이는 다른 오스트랄로피테신과 유사한 수준이다.[21] 뇌 구조는 대체로 오스트랄로피테신과 비슷하지만, 안와전두피질 영역은 현생 인류와 더 유사한 특징을 보인다.[13]
전체적으로 ''A. 세디바''의 두개골 형태는 ''A. 아프리카누스''와 가장 유사하다. 그러나 MH1은 ''A. 아프리카누스''에 비해 머리가 더 작고, 두개골 천장이 가로로 더 넓으며, 두정골 벽이 더 수직에 가깝게 기울어져 있고, 측두선 사이의 간격이 더 넓다. 호모 속과 유사한 특징으로는 덜 발달된 눈썹 능선, 덜 돌출된 광대뼈, 덜 튀어나온 얼굴(낮은 전돌증), 그리고 약간의 턱이 있다는 점 등이 있다.[4] 하지만 인류학자 윌리엄 킴벨(William Kimbel)과 요엘 락(Yoel Rak)은 이러한 ''호모'' 속과 유사한 특징들이 스테르크폰테인 유적 4번 지층에서 발견된 일부 ''A. 아프리카누스'' 두개골에서도 나타나며, 이는 어린 개체의 특징으로 성숙하면서 사라졌을 가능성이 있다고 지적했다. 또한, 얼굴의 돌출 정도(전돌증)를 코의 가장 아랫부분이 아닌 전비극을 기준으로 측정하면 MH1은 ''A. 아프리카누스''의 범위 안에 속한다고 주장했다.[14] 치아는 오스트랄로피테신 중에서는 매우 작은 편으로, 초기 ''호모'' 속의 범위에 해당한다. 그러나 어금니가 입 뒤쪽으로 갈수록 점진적으로 커지고(두 번째 어금니가 가장 큰 ''호모'' 속과 다름) 치아의 융선(cusp)들이 더 가깝게 배치되어 있다는 점은 ''호모'' 속과 다른 특징이다.[4]
턱뼈 아래쪽 가지(하악골 ramus)의 모양은 MH1과 MH2 사이에서 상당한 차이를 보인다. MH1의 것은 더 높고 넓으며, 앞과 뒤 경계가 거의 수직이고 평행하다. 반면 MH2는 앞 경계가 오목하고 경계선이 평행하지 않다. 또한 MH1의 하악골의 근돌기는 뒤쪽으로 기울어져 있고 하악 절흔이 깊고 비대칭적인 반면, MH2는 기울어지지 않은 근돌기와 얕은 하악 절흔을 가지고 있다. 현대 유인원에서 관찰되는 패턴과 비교할 때, 이러한 뚜렷한 차이는 성적 이형성이나 MH1이 어린 개체라는 점만으로는 설명하기 어려울 정도로 크다. 이는 ''A. 세디바''가 개체 간 형태적 다양성이 매우 큰 종이었을 가능성을 시사한다.[15]
3. 2. 몸통
MH1은 4개의 경추, 6개의 흉추, 2개의 요추를 보존하고 있으며, MH2는 2개의 경추, 7개의 흉추, 2개의 요추, 1개의 천골을 보존하고 있다.[19] 목 척추의 요추전만(lordosis, 인간형 곡선)과 관절은 인간과 유사한 머리 자세를 가졌음을 시사한다. 그러나 목 척추의 전반적인 해부학적 구조는 유인원과 유사하여 훨씬 더 뻣뻣한 목을 가졌을 것으로 보인다. ''A. 세디바''는 인간에게서 발견되는 상완 신경총이 없었다. 인간의 상완 신경총은 팔과 손의 신경 및 근육 지배를 통해 운동 제어를 향상시키는 역할을 한다.[18]인간과 마찬가지로, ''A. 세디바''는 비인간 유인원의 6개 척추와 달리 5개의 척추로 구성된 유연한 허리뼈를 가지고 있었고, 습관적인 직립 보행과 일치하는 요추전만을 보였다. 그러나 ''A. 세디바''는 매우 유연한 허리뼈와 과장된 요추전만을 가지고 있었는데,[19] 이는 ''A. 세디바''에서 추정되는 과내전(overpronation) 보행 시 발생하는 안쪽으로 향하는 토크를 상쇄하는 데 관여했을 수 있다.[31]
MH1은 2개의 상부 흉추 갈비뼈, 1개의 중간 흉추 갈비뼈, 3개의 하부 흉추 갈비뼈를 보존하고 있으며, MH2는 4개의 연속적인 상부에서 중간 흉추 갈비뼈와 3개의 하부 흉추 갈비뼈가 척추와 연결되어 있다.[19] 이는 ''A. 세디바''가 유인원처럼 좁은 상체를 가졌음을 시사하지만, 골반의 인간과 유사한 해부학적 구조는 ''A. 세디바''가 넓고 인간과 같은 하부 흉곽을 가졌을 가능성을 시사한다. 좁은 상체는 보행 시 팔 흔들기를 방해하고 흉곽 용적을 제한하여, 격렬한 호흡이나 빠른 보행, 장거리 달리기에 불리했을 것이다. 반면, ''A. 세디바''는 인간처럼 좁은 허리, 재배치된 복사근, 그리고 뒤쪽의 더 넓은 장늑근을 가지고 있었다. 이러한 특징들은 몸통의 측면 굴곡을 상쇄하여 보행 효율을 높이는 데 기여했을 것이다.[20]
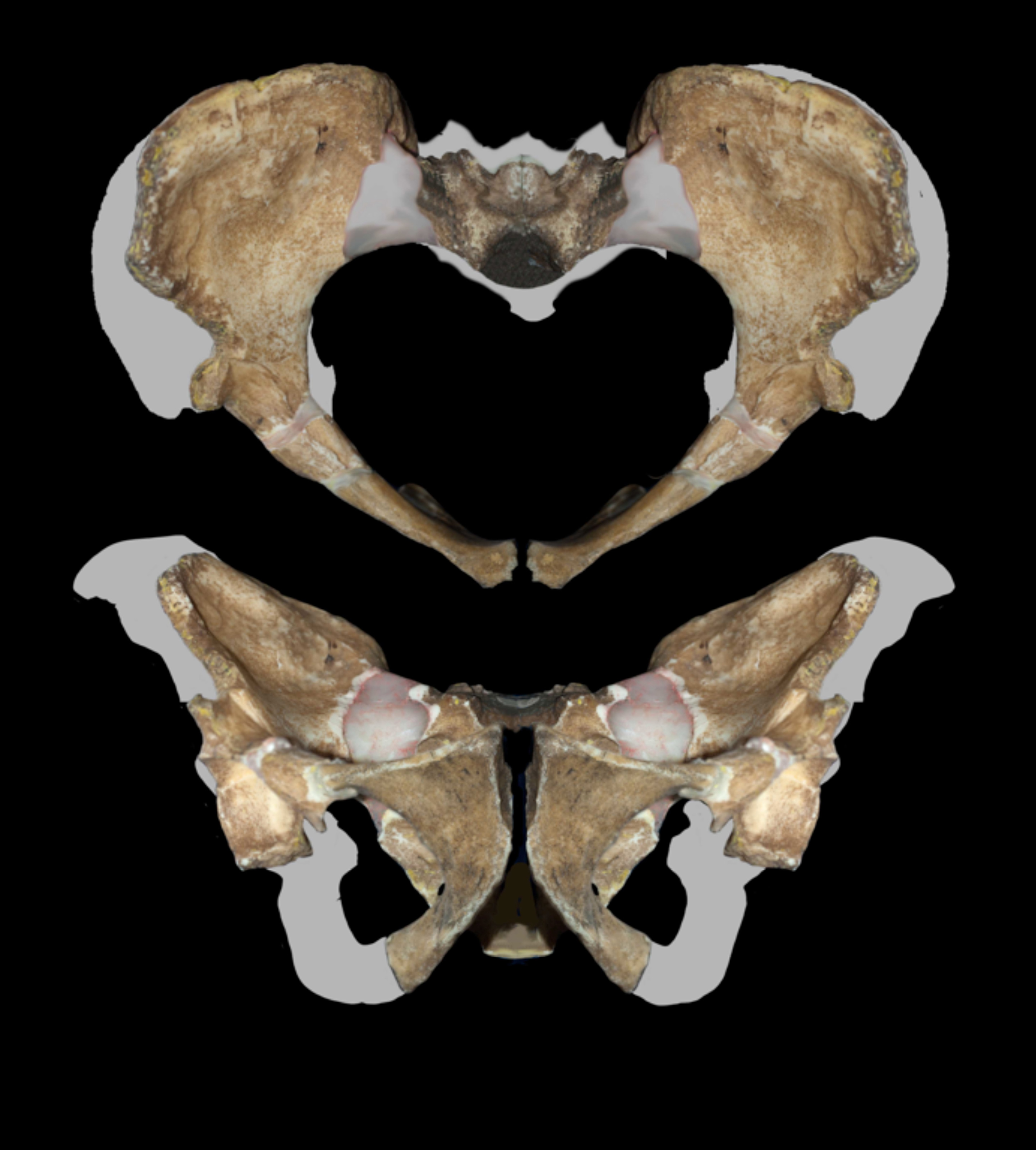
골반은 초기 ''호모'' 속 및 ''H. ergaster''와 여러 특징을 공유한다. 또한 케냐 코오비 포라(Koobi Fora)에서 발견된 KNM-ER 3228, 탄자니아 올두바이 협곡(Olduvai Gorge)의 OH 28과도 유사한 특징을 보이는데, 이 화석들은 특정 종으로 분류되지 않고 일반적으로 ''호모'' 속(spp.)으로 분류된다.[4] 비구(acetabulum)와 천골(sacrum)을 따라 엉덩이 신전(hip extension)을 개선하는 구조가 더 발달했고, 장골대퇴인대(iliofemoral ligament) 부착 부위가 확대되어 엉덩이의 순간 회전 중심(instantaneous center of rotation) 뒤로 체중을 이동시키는 데 유리했으며, 비구와 장골 날개(iliac blade)를 따라 교대하는 골반 기울기(pelvic tilt)를 조절하는 구조가 더 발달했고, 비구와 좌골 결절(ischial tuberosity) 사이의 거리가 더 멀어 햄스트링(hamstring)의 모멘트 팔(moment arm)을 줄였다. 이는 ''A. 세디바''가 인간과 유사한 보폭으로 걸을 수 있게 했을 가능성이 있다. 엉덩이 관절은 ''H. habilis'' 표본 OH 62보다 인간에 더 가까운 하중 지지 패턴을 보였던 것으로 추정된다.[4]
''A. 세디바''의 산도(birth canal)는 다른 오스트랄로피테쿠스 속보다 부인형 골반(gynecoid pelvis, 일반적인 여성형 골반)에 가까운 형태를 보이며, 다른 오스트랄로피테쿠스 속은 더 넓은 편평 골반(platypelloid pelvis) 형태를 가진다. 하지만 ''A. 세디바''는 신생아의 뇌 크기(따라서 머리 크기)가 작았기 때문에 완전히 부인형 골반은 아니었다. 인간과 마찬가지로 산도는 시상면(sagittal plane, 앞에서 뒤로) 직경이 증가했고, 치골(pubis)이 위로 향하는 형태를 보였다.[21]
3. 3. 상지(팔)
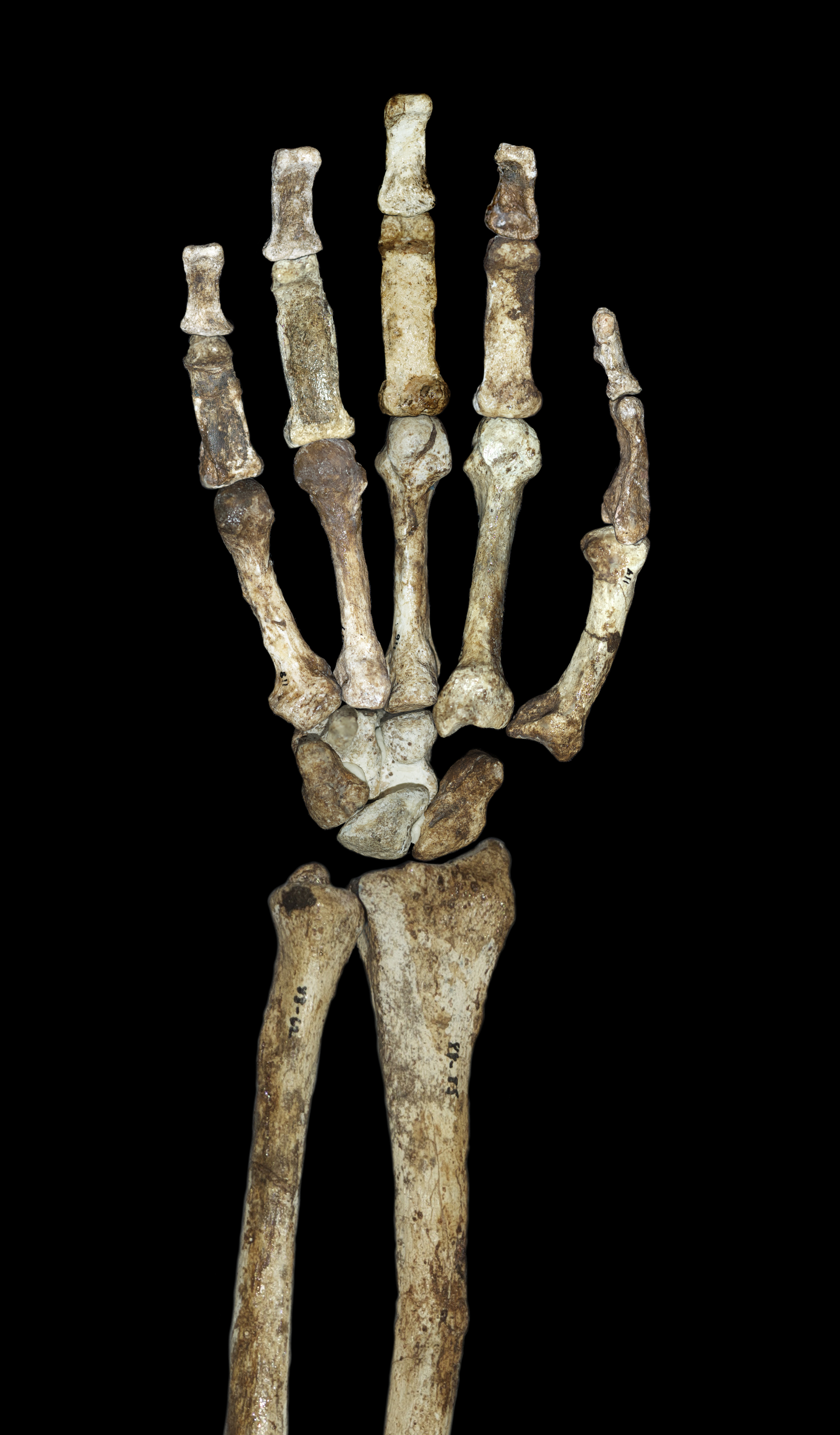
다른 오스트랄로피테쿠스 및 초기 ''호모''와 마찬가지로, ''A. 세디바''는 상대적으로 긴 팔, 84의 높은 상완 지수(전완 대 상완골 비율) 및 큰 관절 표면을 가진 다소 유인원과 같은 상체 비율을 가지고 있었다. 오스트랄로피테쿠스의 유인원과 같은 상지 구성이 수목 생활을 나타내는 것인지, 아니면 더 인간과 같은 팔 해부학을 채택하려는 주요 선택적 압력이 없는 상태에서 유인원 공통 조상으로부터 상속된 기본 특성인지에 대한 논쟁이 있다. 어깨는 으쓱하는 자세를 하고 있으며, 어깨뼈는 잘 발달된 겨드랑이 경계를 가지고 있으며, 원뿔 결절(어깨 관절 주변의 근육 부착에 중요)이 잘 정의되어 있다.[4] 쇄골의 근육 흉터 패턴은 인간과 같은 운동 범위를 나타낸다. 어깨뼈는 오랑우탄과 관절와의 크기(어깨 관절을 형성)와 척추와의 각도 면에서 가장 유사하지만, 어깨뼈의 모양은 인간과 침팬지와 가장 유사하다. 상완골은 인간 및 아프리카 유인원과 달리 비틀림 정도가 낮으며(짧은 쇄골과 함께) 이는 어깨뼈가 다른 오스트랄로피테쿠스처럼 등에 더 높이 위치하지만 ''호모''와 유사하게 정중선에서 더 멀리 떨어져 있음을 시사한다.[22] 팔의 유인원과 같은 특성은 더 고대 ''A. 아파렌시스''보다 ''A. 세디바''에서 더욱 두드러지며, ''A. 아파렌시스''가 ''A. 세디바''의 조상이라면 이는 수목 생활로의 적응 변화를 나타낼 수 있다.[23]
팔꿈치 관절에서, 상완골의 외측 및 내측 상과는 다른 오스트랄로피테쿠스 및 비인간 아프리카 유인원과 매우 유사하게 길어진다. 상완골은 또한 전완을 굽히는 상완요골근을 지지하기 위해 팔꿈치 관절에 발달된 능선을 가지고 있다. 비인간 아프리카 유인원과 마찬가지로 요골에 이두근이 강하게 부착되고 척골에 삼두근이 강하게 부착된다. 그러나 이두근과 상완근의 기계적 이점은 적다.[22] 척골은 또한 척측수근굴근에 대한 강한 부착을 지원한다. 주두와 오목은 크고 깊으며, 팔이 펴져 있는 동안 팔의 안정성을 유지하는 데 중요한 두드러진 도르래 용골이 있다. 손가락 뼈는 길고 튼튼하며 구부러져 있으며, 손가락을 굽히는 데 중요한 강한 천지굴근을 지지한다.[4] 이는 때때로 오스트랄로피테쿠스의 수목 생활 행동의 증거로 주장된다. 손은 또한 ''호모''와 매우 유사한 상대적으로 긴 엄지손가락과 짧은 손가락을 특징으로 하며, 이는 복잡한 석기를 만들고 사용하는 데 중요한 정밀 파지를 시사할 수 있다.[24]
3. 4. 하지(다리)
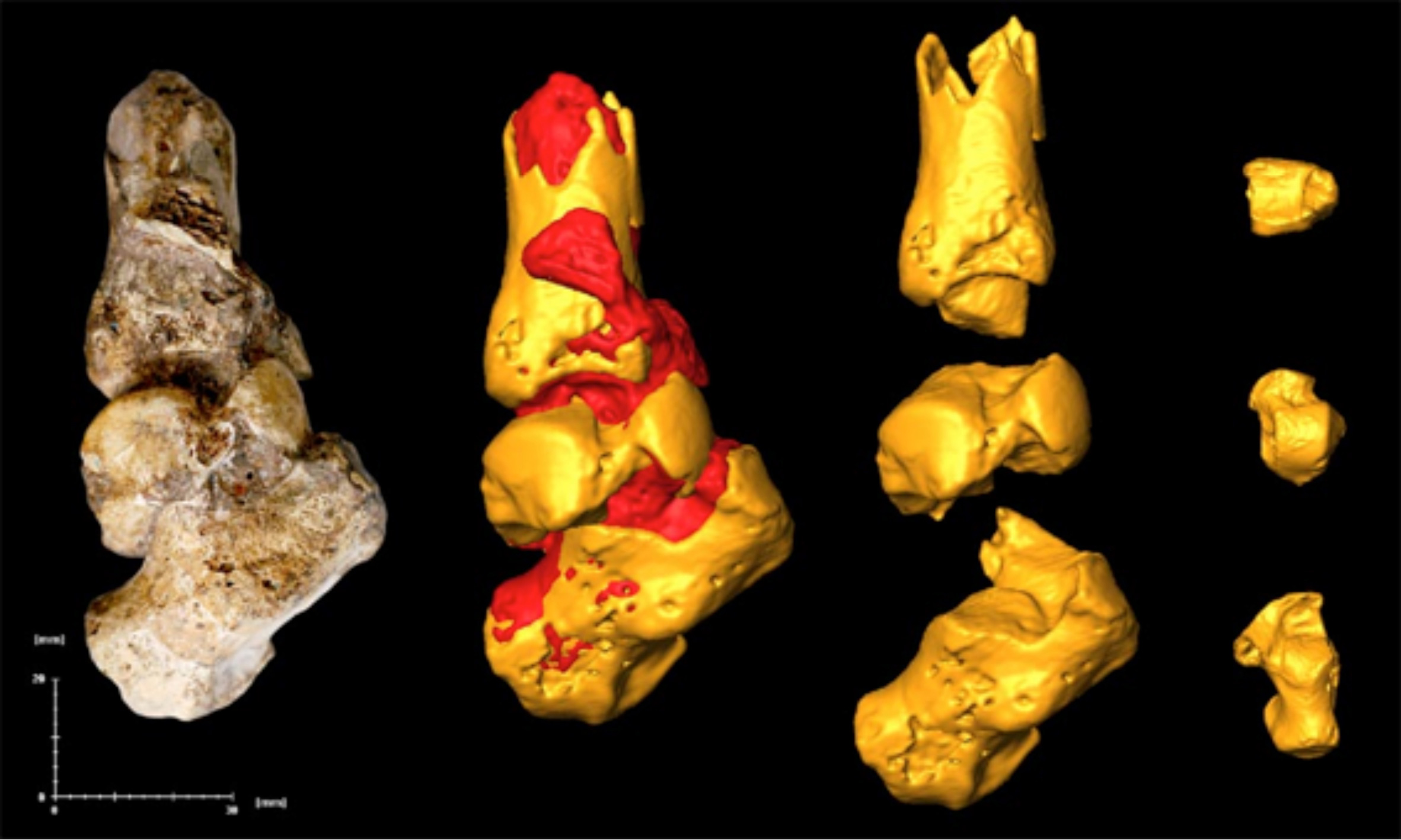
다른 오스트랄로피테쿠스처럼, 발목, 무릎, 엉덩이 관절은 습관적인 두 발 보행을 나타낸다. 다리 뼈는 ''A. afarensis''의 다리 뼈와 매우 유사하다. 발목은 대체로 사람과 유사하며, 인간과 유사한 아킬레스건을 가졌을 가능성이 있다.[26]
목말뼈는 튼튼하며 사람 외의 유인원과 더 유사한 특징을 보이는데, 안쪽으로 꼬인 목과 낮은 목 비틀림 각도가 특징이다. ''A. sediba''가 사람과 같은 발 아치를 가졌는지, 아니면 발이 유인원과 더 유사했는지에 대해서는 논쟁이 있다.[25] 발꿈치뼈는 45도 각도로 기울어져 있으며, 앞쪽에서 뒤쪽으로 두드러지게 기울어져 있고, 특히 종골 활차에서 가장 강하게 나타난다. 튼튼한 종골 활차는 종아리에서 발목까지 이어지는 강한 종아리 근육이 있었음을 시사한다. 발에는 인간과 ''A. afarensis''에게서 보이는 외측 저측 결절(정상적인 인간 보행 시 발꿈치가 땅에 닿을 때 힘을 분산시키는 데 관여할 수 있는 부분)이 없다.[4][26] 발꿈치뼈의 가늘고 튼튼한 복사뼈(발목 양쪽에 있는 뼈 돌출부)는 유인원과 매우 유사하며, 이는 발꿈치뼈와 목말뼈 사이의 힘 전달 효율이 낮고, 발 중간 부위에서 유인원과 같은 이동성을 보였음을 의미한다. ''A. sediba''의 발은 고릴라에서 보이는 상태와 가장 유사하며, 기능적으로는 ''A. africanus''의 발과 동일했을 수 있다.[26][27]
4. 고생물학
2010년, 리 버거(Lee Berger)와 동료 연구자들은 MH1을 모식 표본으로, MH2를 부모식 표본으로 지정하여 ''오스트랄로피테쿠스 세디바''(''Australopithecus sediba'')라는 새로운 호미닌 종을 공식적으로 기술했다. 종명 sediba|세디바st는 남아프리카 공화국 현지 세소토어로 "샘" 또는 "우물"을 의미한다.[4]
발견자들은 ''A. 세디바''가 특히 골반과 다리에서 후대의 호모 에르가스터나 H. 에렉투스와 많은 특징을 공유한다는 점에 주목했다. 치아 특징 역시 호모 속과의 유사성을 보였다. 이러한 이유로 그들은 ''A. 세디바''가 오스트랄로피테쿠스 속과 호모 속 사이의 전이 화석일 수 있다고 제안했다.[4][5]
하지만 화석이 발견된 지층의 연대는 195만 년 전에서 178만 년 전 사이로 측정되었는데, 이는 당시 알려진 가장 오래된 ''호모'' 속 화석인 에티오피아 하다르 지역의 ''H. 하빌리스''(약 233만 년 전)보다 훨씬 이후의 시대이다.[4] 현재 가장 오래된 ''호모'' 속 표본은 에티오피아 레디-게라루 지역에서 발견된 약 280만~275만 년 전의 LD 350-1 턱뼈 화석이다.[6] 이 연대 차이를 설명하기 위해, 발견자들은 ''A. 세디바''가 같은 지역에 더 이른 시기에 살았던 오스트랄로피테쿠스 아프리카누스 집단에서 진화했으며, 이후 ''호모'' 속이 ''A. 세디바''에서 갈라져 나왔을 것이라는 가설을 제시했다.[4] 이 가설은 ''A. 아프리카누스''와 ''A. 세디바'' 사이에 약 80만 년의 유령 계통(화석 기록이 없는 가상의 계통)이 존재함을 암시한다.[14] 또한, 2011년에 말라파 호미닌의 연대가 약 198만 년 전으로 더 정확히 측정되면서, 이는 당시 알려진 가장 오래된 ''H. 에르가스터''/''H. 에렉투스''보다 앞서는 시기였기 때문에, ''A. 세디바''가 초기 ''호모'' 속인 ''H. 하빌리스''나 ''H. rudolfensis'' 대신 ''H. 에르가스터''/''H. 에렉투스''의 직접적인 조상일 수 있다는 주장도 제기되었다.[2] 현재 ''A. 세디바''는 인류의 요람 지역에서 ''H. 에르가스터''/''H. 에렉투스'' 및 파란트로푸스 로부스투스와 같은 다른 호미닌 종들과 동시대에 살았던 것으로 여겨진다.[7]
다른 한편으로, ''A. 세디바''가 ''호모'' 속과는 별개로, ''A. 아프리카누스''의 후기 생존 형태이거나 자매 종일 가능성도 제기된다. 이 경우, ''A. 세디바''가 보이는 ''호모'' 속과 유사한 특징들은 독립적으로 진화한 결과(동형성)로 해석될 수 있다.[14][8][9][10][11] 초기 ''호모'' 속의 화석 기록은 매우 부족하고 단편적이어서, 종들 간의 해부학적 비교와 계통 관계 설정에 어려움이 많다.[11] 실제로 ''A. 아프리카누스'', ''A. afarensis'', ''A. garhi'' 등 다른 오스트랄로피테쿠스 종들도 ''호모'' 속의 조상 후보로 제안되어 왔으며, 이 문제는 여전히 고인류학계의 주요 논쟁거리이다.[6] 또한, 모식 표본인 MH1이 어린 개체라는 점도 해석의 어려움을 더한다. 일부 학자들은 MH1에서 보이는 ''호모'' 속과 유사한 얼굴 특징 중 일부는 성체가 되면서 사라졌을 수도 있다고 지적한다.[14]
현재 오스트랄로피테신의 분류는 다소 혼란스러운 상태이다. ''오스트랄로피테쿠스'' 속은 명확한 단계통군이라기보다는 비슷한 생리적 특징을 공유하는 여러 호미닌 종들을 묶어놓은 진화 등급으로 간주되기도 하며, 대부분의 경우 특정 종이 다른 종과 구체적으로 어떤 관계에 있는지 명확히 밝혀지지 않았다.[12]
4. 1. 식단
식물 규소체(미세한 식물 잔해) 분석, 두 표본의 치태 및 탄소 동위 원소 분석 결과는 ''오스트랄로피테쿠스 세디바''가 주로 C3 숲 식물을 먹었음을 보여준다. 이는 주변 사바나 환경에 C4 식물이 널리 퍼져 있었음에도 불구하고 나타난 특징이다. 이러한 식습관은 현대의 사바나 침팬지에게서도 발견되며, 플리오세 초기의 ''아르디피테쿠스 라미두스''에게서도 비슷한 식단이 추정된다. 하지만 이는 다른 초기 호미닌들과는 상당히 다른 모습이다.MH1 개체의 치아 두 개에서 총 38개의 식물 규소체가 발견되었는데, 이 중 15개는 쌍떡잎식물, 9개는 외떡잎식물과 일치했고, 나머지 14개는 종류를 특정하기 어려웠다. 외떡잎식물 규소체는 물이 풍부하고 그늘진 곳에서 자라는 C3 풀이나 사초에서 온 것으로 보이며, 나머지 규소체들은 과일, 잎, 나무 또는 나무껍질에서 유래한 것으로 추정된다. 나무껍질은 단백질과 당분 함량이 높아 다른 영장류가 즐겨 먹지만, 이를 규칙적으로 먹었던 다른 호미닌은 아직까지 알려지지 않았다.
치아의 미세한 마모 흔적을 분석한 결과, 말라파에서 발견된 두 호미닌 개체(MH1, MH2)는 단단한 음식을 먹었음을 알 수 있다. 이 마모의 복잡성 정도는 ''호모 에렉투스''와 ''파란트로푸스 로부스투스''의 중간 수준에 해당한다.[28] 그럼에도 불구하고 ''오스트랄로피테쿠스 세디바''의 턱 구조는 다른 초기 호미닌에 비해 강한 힘을 내는 데 그다지 적합하지 않은 것으로 보인다. 이는 ''오스트랄로피테쿠스 세디바''가 단단하거나 질긴 음식을 처리하는 능력에 크게 의존하지 않았을 가능성을 시사한다.[29][30]
''오스트랄로피테쿠스 세디바''가 C3 숲 식물을 먹는 일반주의자 초식 동물이었다는 해석은 이들이 어느 정도 나무 위에서 생활했다는 점과도 일치한다. 나무껍질처럼 부수기 어려운 음식까지 먹을 수 있었던 ''오스트랄로피테쿠스 세디바''는 주로 과일만 먹는 현대의 사바나 침팬지보다 훨씬 좁은 활동 범위를 가졌으리라 추정된다.[28]
4. 2. 보행
다른 오스트랄로피테쿠스처럼, 발목, 무릎, 엉덩이 관절은 습관적인 두 발 보행을 나타낸다. 다리 뼈는 ''A. afarensis''의 다리 뼈와 매우 유사하다. 발목은 대체로 사람과 유사하며, 인간과 유사한 아킬레스건을 가졌을 수도 있다.[26]
목말뼈는 튼튼하고 사람 외의 유인원과 더 유사하며, 안쪽으로 꼬인 목과 낮은 목 비틀림 각도를 특징으로 한다. ''A. sediba''가 사람과 같은 발 아치를 가졌는지, 아니면 발이 유인원과 더 유사했는지에 대해서는 논쟁이 있다.[25] 발꿈치뼈는 45도 각도로 기울어져 있으며, 앞쪽에서 뒤쪽으로 두드러지게 기울어져 있고, 종골 활차에서 가장 강하게 나타난다. 튼튼한 종골 활차는 종아리에서 발목까지 뻗어 있는 강한 종아리 근육을 나타낸다. 발에는 인간과 ''A. afarensis''에서 볼 수 있는 외측 저측 결절(정상적인 인간 보행에서 발꿈치가 땅에 닿을 때 힘을 분산시키는 데 관여할 수 있음)이 없다.[4][26] 발꿈치뼈의 가늘고 튼튼한 복사뼈 (발목 양쪽에 있는 뼈의 돌출부)는 유인원과 매우 유사하며, 발꿈치뼈와 목말뼈 사이의 힘 전달이 덜 효율적이고, 발 중간 부위에서 유인원과 같은 이동성을 보인다. ''A. sediba''는 고릴라에서 볼 수 있는 상태와 가장 유사하며, 발은 ''A. africanus''의 발과 기능적으로 동일했을 수 있다.[26][27]
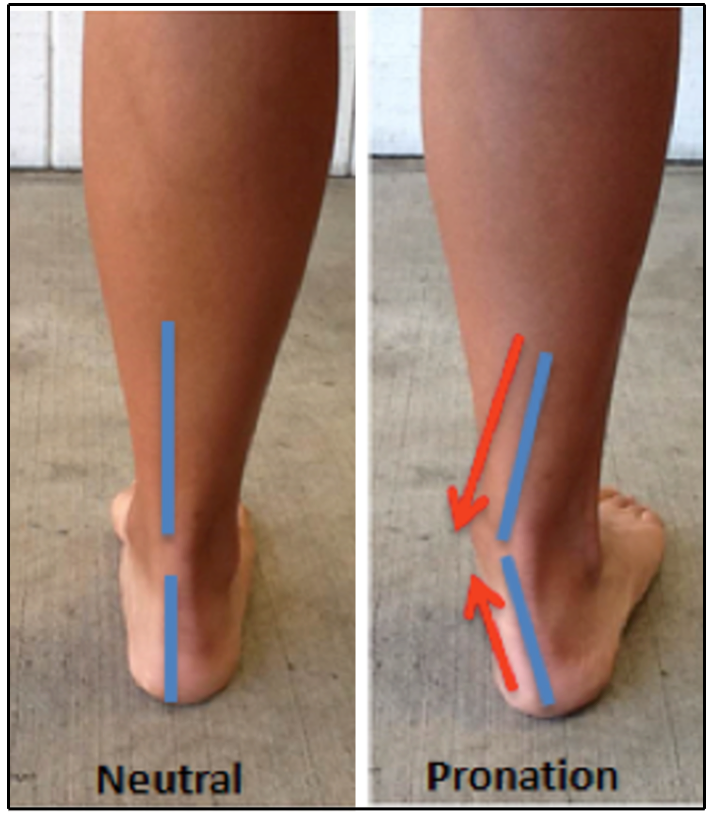
걸을 때, ''오스트랄로피테쿠스 세디바''는 서는 동안 체중이 과장되게 안쪽으로 이동하는 발목 관절의 과내전을 보였을 수 있다. 현대 인간의 과내전자의 경우, 발은 스윙 단계에서 심하게 내전되며, 땅과의 접촉은 발의 바깥쪽 가장자리에서 먼저 이루어져 전체 다리를 안쪽으로 회전시키는 높은 토크를 발생시킨다. 이와 유사하게, ''오스트랄로피테쿠스 세디바''의 대퇴직근과 넙다리두갈래근의 부착점은 다리, 엉덩이 및 무릎을 가로지르는 중앙선 방향의 스트레스와 일치한다. 이러한 걷는 방식은 현대 인간 해부학에는 이상적이지 않으며, 과내전자는 족저근막염, 정강이 부목, 그리고 경골 피로 골절이 발생할 위험이 더 높다. 이를 상쇄하기 위해, ''오스트랄로피테쿠스 세디바''는 딱딱한 인간형 발 중앙부 대신 이동 가능한 발 중앙부를 사용했을 수 있으며, 이는 발목에 과도한 스트레스를 가하는 것을 막았을 수 있다.[31]
과내전 보행과 관련된 일련의 적응은 다른 호미닌에서는 확인되지 않았으며, ''오스트랄로피테쿠스 세디바''가 왜 이것을 발달시켰는지는 불분명하다.[31] 이동 가능한 발 중앙부는 광범위한 등반 행동에도 유용했을 것이므로,[4][31][26] 과내전은 습관적인 이족 보행과 수목 생활 사이의 타협점이었을 수 있다.[31]
4. 3. 출산
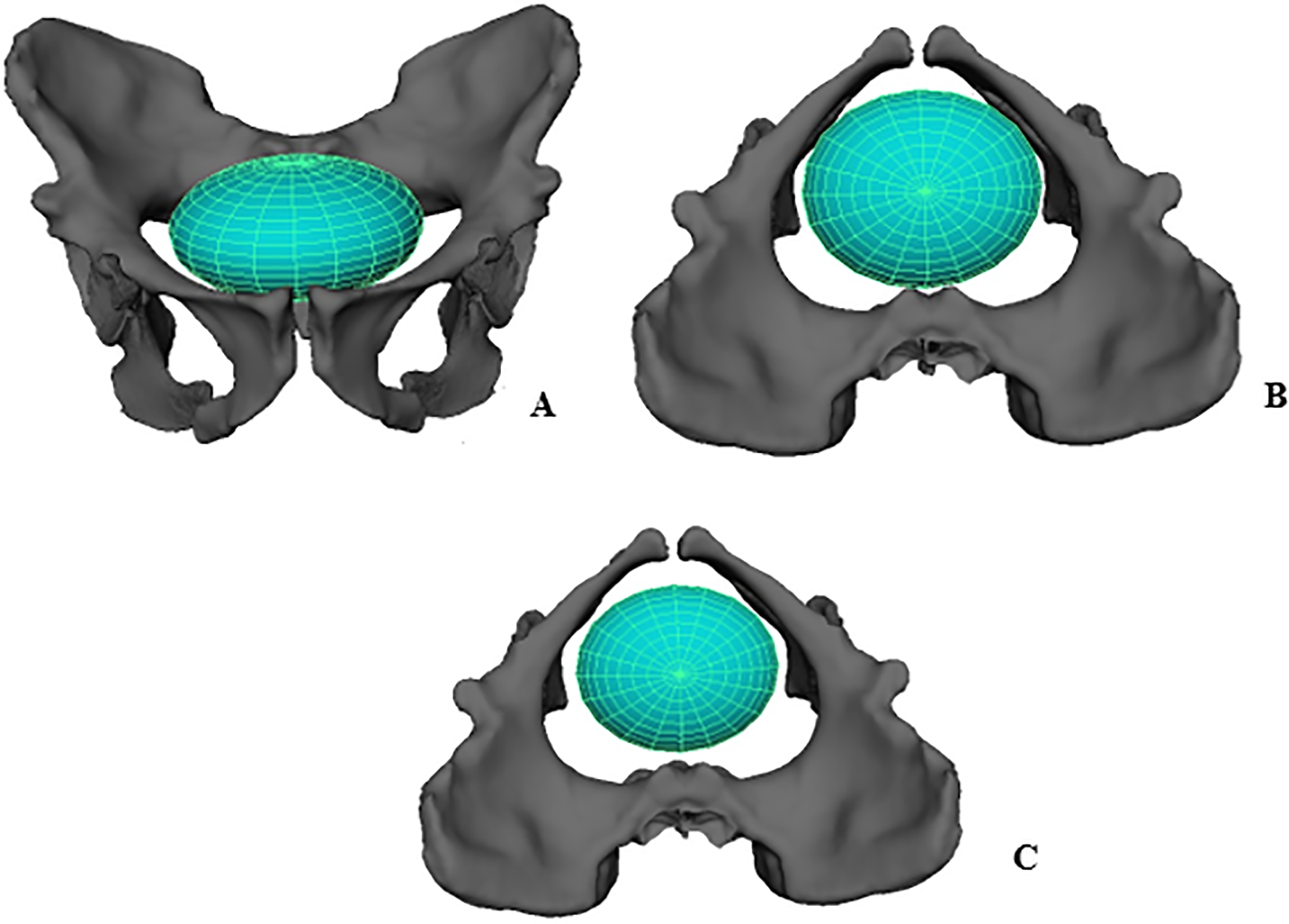
암컷 ''A. 세디바''의 골반 입구 크기는 시상 방향 80.8mm, 가로 방향 112.4mm로 추정된다. 신생아 머리의 가장 긴 부분이 89.2mm로 추정되므로, 신생아는 다른 호미닌과 비슷하게 골반 입구에 가로 방향으로 들어갔을 가능성이 높다. 골반 입구의 중간 평면은 최소 96.9mm까지 좁아지기 때문에, 신생아가 태어나는 동안 회전할 필요가 없었을 수도 있다. 다만, 골반 입구 치수는 어린 수컷의 좌골을 포함하여 복합적으로 재구성하여 계산되었으므로, 실제 산도는 계산된 것보다 더 컸을 가능성도 있다. 신생아의 어깨 너비는 74.3mm로 추정되어 머리보다 출산을 더 방해하지는 않았을 것이다. 따라서 신생아는 가장 좁은 지점에서 산도의 약 92.1%를 차지하게 되는데, 이는 인간이 아닌 유인원이나 다른 오스트랄로피테쿠스에서처럼 완전히 회전하지 않는 출산에 충분한 공간을 제공한다(부분적인 회전 가능성도 제기됨). 회전 없이 통과하는 것이 가능하지만, 중간 평면은 앞뒤 방향으로 확장되어 있어, 머리의 가장 긴 부분이 이 확장에 맞춰 정렬되도록 회전했다면 신생아를 위한 공간이 더 확보되었을 것이다.[32]
이에 비해 현대 인류는 신생아의 완전한 회전이 필요한 훨씬 더 어렵고 복잡한 출산을 한다. 이는 인간 신생아의 큰 뇌와 그에 따른 머리 크기, 그리고 뻣뻣한 어깨가 산도를 통과하기 어렵게 만들기 때문이다. ''A. 세디바'' 신생아의 뇌 크기를 145.8–180.4 cc로 추정하면, 신생아 머리 크기는 침팬지 신생아와 비슷한 73mm x 89mm였을 것으로 보인다.[32]
4. 4. 성장
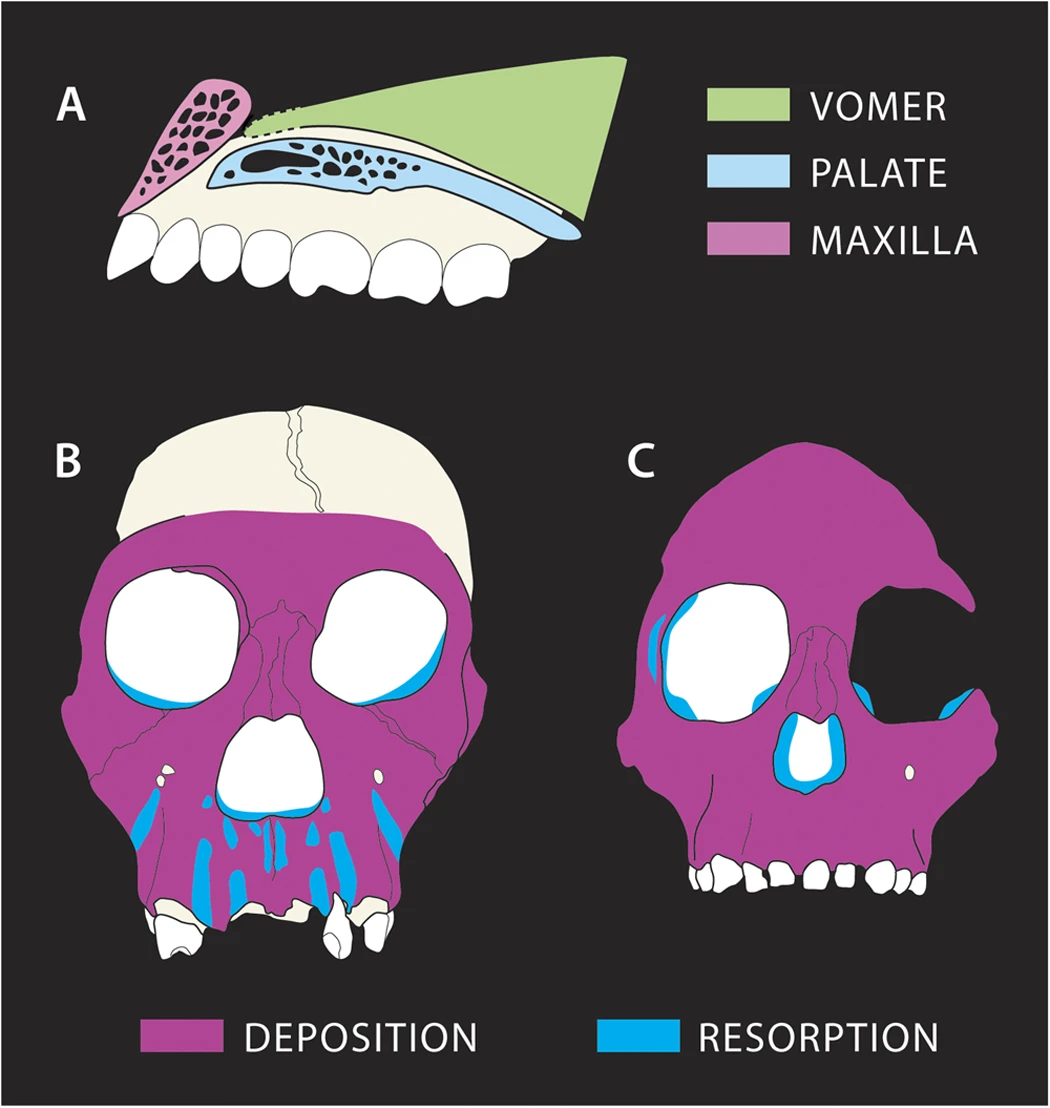
현대 영장류의 성체와 신생아 뇌 크기 사이의 경향을 이용하여 ''오스트랄로피테쿠스 세디바'' 신생아의 뇌 크기는 153ml에서 201ml 사이였을 것으로 추정된다. 이는 다른 오스트랄로피테신에서 추정된 크기와 유사하다.[21] 성체가 된 MH1 개체의 뇌 부피는 약 420ml~440ml로 추정되는데, 이는 오스트랄로피테신의 전형적인 뇌 크기 범위에 속한다.[4]
MH1 개체의 성장 궤적은 다른 호미닌과 비교했을 때 눈에 띄게 달랐던 것으로 보인다. 코에서 윗입술까지 이어지는 뼈인 비상악 복합체에서 상당한 뼈 흡수가 일어났는데, 특히 앞니의 치근 부위에서 두드러졌다. 이는 나이가 들면서 얼굴이 점점 더 돌출되는 경향을 보이는 ''A. africanus''나 ''A. afarensis''와는 대조적인 특징이다. ''P. robustus'' 역시 위턱뼈 흡수 현상을 보이지만, MH1의 흡수는 앞니를 따라 볼뼈 근처의 견치와까지 확장되어 ''P. robustus''의 평평한 얼굴과는 다른, 다소 튀어나온 중안면 형태를 만들었다. 볼뼈에 매우 가깝게 뼈 흡수가 발생했기 때문에, MH1이 ''A. africanus''의 특징인 튀어나온 광대뼈를 가지지 않은 이유를 설명할 수 있다. MH1은 이미 모든 영구치가 난 상태였으므로, 치아 맹출이 아래쪽 얼굴의 리모델링에 영향을 미치지는 않았을 것이다. 그럼에도 불구하고 작은 어금니 크기는 중안면 형성에 영향을 주었을 수 있다.
''A. sediba''는 일반적인 초기 호미닌의 식단과는 상당히 다른 식단을 가졌을 것으로 추정되는데, 주로 어린 잎을 먹는 현생 올리브콜로부스 원숭이의 식단과 유사했을 가능성이 있다. 흥미롭게도 ''A. sediba''와 올리브콜로부스 원숭이는 얼굴 뼈 성장 패턴이 유사한 것으로 보인다. 이는 다른 호미닌과 다른 턱 사용 방식(턱 로딩 패턴)을 반영하여, ''A. sediba''에서 뼈의 흡수와 축적 패턴이 달랐음을 시사할 수 있다. 예를 들어, MH1의 눈구멍 가장자리는 굽어 있지만, ''A. africanus''에서는 움푹 들어가 있는데, 이는 ''A. africanus''에서 뼈 흡수가 일어나는 부위에 ''A. sediba''에서는 뼈가 축적되었음을 나타낼 수 있다.[33]
또한, MH1과 MH2 개체 사이에서 하악골 형태에 상당한 차이가 발견된다. MH1의 하악골 가지(ramus)는 더 높고 넓으며, 앞뒤 경계가 거의 수직이고 평행하다. 반면 MH2는 앞 경계가 오목하고 경계선이 평행하지 않다. MH1의 근돌기는 뒤로 기울어져 있고 하악 절흔이 깊고 비대칭적인 반면, MH2는 얕은 하악 절흔과 함께 휘지 않은 근돌기를 가지고 있다. 이러한 현저한 차이는 현대 유인원에서 관찰되는 성적 이형성이나 MH1의 어린 개체 상태만으로는 설명하기 어려울 정도로 크다. 이는 ''A. sediba''가 골격적으로 매우 다양한 특징을 가진 종이었을 가능성을 시사한다.[15]
4. 5. 병리학
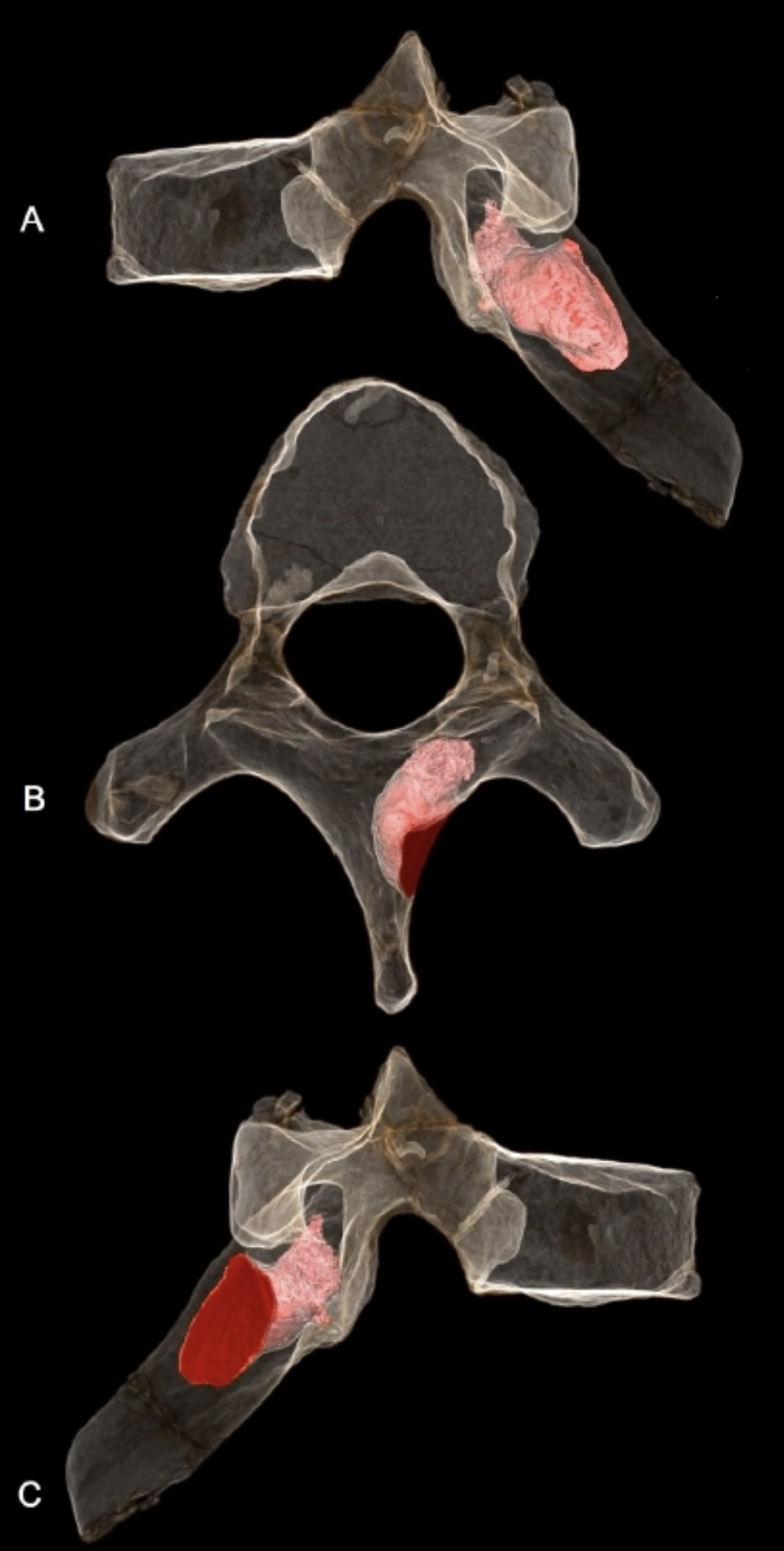
MH1 개체의 여섯 번째 흉추 오른쪽 판에서는 침투성 뼈 종양이 발견되었는데, 이는 아마도 양성 골성 골종으로 추정된다. 이 병변은 6.7mm 깊이로 뼈에 침투해 있었고 너비는 5.9mm였으며, 사망 당시에도 활성 상태였던 것으로 보인다. 종양이 척수관을 관통하지는 않아서 신경학적 합병증을 유발하지는 않았을 것으로 보이며, 척추 측만증(척추의 비정상적인 굽음)의 증거도 없다. 하지만 이 종양은 어깨뼈와 등의 오른쪽 위쪽 사분면 움직임에 영향을 미쳐 급성 또는 만성 통증, 근육 장애 또는 근육 경련을 유발했을 수 있다. ''오스트랄로피테쿠스 세디바''가 나무를 오르는 능력이 필요했을 수 있다는 점을 고려하면, 승모근, 척추기립근, 능형근과 같은 근육들의 부착 지점 근처에 병변이 위치하여 정상적인 움직임 패턴을 제한했을 가능성이 있다. MH1은 인류 화석 기록상 암이 진단된 최초의 사례로, 기존에 가장 오래된 것으로 알려졌던 남아프리카 공화국 스와르트크란스에서 발견된 180만 년에서 160만 년 전의 골육종을 나타내는 SK 7923 중족골 조각보다 최소 20만 년 앞선다. 영장류에서 종양은 일반적으로 발병률이 낮기 때문에 인류 화석 기록에서도 드물게 발견되며, 초기 인류 역시 현대 영장류와 비슷한 발병률을 가졌을 가능성이 높다. 어린 개체였던 MH1에서 뼈 종양이 발견된 것은, 뼈 종양이 주로 젊은 개체에게 발생하는 일반적인 경향과 일치한다.[34]
MH1과 MH2 개체 모두 둔력 외상과 일치하는 사망 시점 전후(perimortem)의 뼈 부상을 가지고 있다. 이는 이들이 발견된 말라파 유적지가 동물들이 우연히 빠지는 자연적인 죽음의 함정 역할을 하는 높은 수직 통로의 바닥이었다는 해석과 잘 들어맞는다. MH1과 MH2는 약 5m 높이에서 떨어졌을 것으로 추정되는데, 바닥에 쌓여 있던 자갈, 모래, 박쥐 구아노 등이 어느 정도 낙하 충격을 완화했을 수 있다. MH1의 경우, 사망 전 골절이 턱뼈와 치아에서 가장 두드러지게 나타나지만, 추락 외에도 낙하하는 물체에 맞아 부상을 입었을 가능성도 배제할 수 없다. MH2는 부상 당시 방어 자세를 취했던 증거를 보이는데, 전완과 손에 하중이 실리고 가슴에 충격이 가해졌으며, 신체의 오른쪽에 사망 전 골절이 확인되었다. 이는 포식이나 명백한 자연사로 인한 사망이 아닌, 외상으로 인한 사망 사례로는 오스트랄로피테쿠스 화석 기록에서 처음으로 확인된 경우이다.[35]
5. 고생태학
말라파 주변의 동굴 네트워크는 약 500 m × 100 m 면적 내에 길고 서로 연결된 동굴 입구들로 구성되어 있다. 말라파 유적지는 최대 30m 깊이에 달하는 동굴 시스템의 바닥 부분이었을 가능성이 있다. 이 동굴들은 북-북동 방향과 북-북서 방향으로 뻗은 단층들의 교차 지점에 위치하며, 호미닌 유해는 북-북서 단층의 3.3 m × 4.4 m × 3.5 m 크기 구획에서 발굴되었다. 이 지층은 20세기 초 석회암 채굴 과정에서 드러났다. 동굴 내부는 물에 의해 운반되어 쌓인 사암으로 이루어진 5개의 퇴적암 상(facies) A부터 E까지로 구성된다. ''A. 세디바''는 상 D에서, 더 많은 호미닌 유해는 상 E에서 발견되었다. MH1과 MH2 개체는 수직으로 최대 40cm 간격을 두고 발견되었다. 상 D는 유석 위에 쌓인 1.5m 두께의 엷은 색 지층이다. 작은 펠로이드가 흔하게 발견되며, 때로는 크고 불규칙한 덩어리로 뭉쳐 있는데, 이는 물에 잠긴 환경에서 퇴적되었음을 시사한다. 펠로이드는 동물의 분변 물질이나 토양 미생물에서 유래했을 수 있다. MH1과 MH2의 보존 상태는 이들이 빠르게 퇴적되었고 거의 이동하지 않았으며, 고결 작용이 일어난 직후 포화대(phreatic, 지하수면 아래) 환경에 놓였음을 나타낸다. 포식 흔적이 없어, 당시 육식동물이 이 장소에 접근하기 어려웠음을 시사한다.[3]
이러한 정황들은 말라파 동굴이 표면에서는 잘 보이지 않는 입구를 가진 '죽음의 덫'이었을 가능성을 제기한다. 동물들은 동굴 입구에서 나는 물 냄새에 유인되었을 수 있으며, 이후 떨어진 동물 사체의 냄새를 맡고 온 육식동물들도 함정에 빠져 죽었을 수 있다. 거대한 암설류(debris flow)가 발생하여 유해들을 지하 시내를 따라 동굴 깊숙한 곳으로 옮겨 퇴적시켰을 것으로 추정되며, 이는 아마도 폭우와 관련이 있을 것이다. 시간이 지나면서 동굴은 결국 붕괴되고 진흙으로 채워졌다.[3]
2010년 기준으로, D 및 E 상에서는 호미닌 외에도 총 209점의 다른 동물 화석이 발견되었다. 확인된 동물군은 다음과 같다.[3][36]
| 분류 | 종 또는 속 |
|---|---|
| 검치호 | 디노펠리스 바를로위 (Dinofelis barlowi) |
| 고양이과 | 표범, 아프리카들고양이, 검은발고양이 |
| 하이에나과 | 점박이하이에나 |
| 개과 | 케이프여우, 아프리카들개 (Lycaon) |
| 몽구스과 | Atilax mesotes, Mungos |
| 사향고양이과 | 사향고양이 |
| 말과 | 말 |
| 멧돼지과 | 돼지 |
| 소과 | 클립스프링어, 메갈로트라구스 (Megalotragus), 큰 알셀라피네 영양, 부시벅, 큰쿠두 |
| 토끼목 | 토끼 |
오늘날 검은발고양이와 케이프여우는 남아프리카의 초원, 덤불, 관목지에 서식하는 고유종이다. 점박이하이에나 역시 건조하고 개방된 서식지에 살며 폐쇄된 숲 환경에서는 거의 발견되지 않는다. 반면, ''디노펠리스''와 ''Atilax''는 일반적으로 폐쇄되고 습한 서식지를 선호했던 것으로 알려져 있다. 이러한 동물상의 조합은 당시 말라파 지역이 약 20km2 범위 내에 폐쇄된 숲 환경과 개방된 초원을 모두 포함하는 모자이크 형태의 서식지였을 가능성을 시사한다.[36]
D 상에서 발견된 육식동물의 분화석(coprolite)에는 ''Podocarpus'' 또는 ''Afrocarpus'' 나무의 꽃가루와 식물규소체, 그리고 미확인 침엽수 및 쌍떡잎식물의 나무 조각이 포함되어 있었다. 풀에서 유래한 식물규소체는 발견되지 않았다. 현재 말라파 유적지는 초원 지대이며, ''Podocarpus''와 ''Afrocarpus'' 나무는 약 30km 떨어진 아프로몬탄 숲 생물군계, 특히 산불이 덜 발생하는 마갈리스버그 산맥의 해발 1500m에서 1900m 사이 협곡에서 발견된다. 이는 초기 플라이스토세 당시 말라파 지역이 오늘날보다 더 시원하고 습했으며, 산불 발생 빈도가 낮아 자연적으로 보호된 지역을 넘어 이러한 숲 식생이 퍼져나갈 수 있었음을 나타낼 수 있다. 또한 당시 말라파는 현재보다 약간 낮은 고도에 위치했을 수 있으며, 계곡과 마갈리스버그 지형이 덜 뚜렷했을 가능성도 있다.[37]
오스트랄로피테쿠스와 초기 ''호모'' 속은 후기 ''호모'' 속보다 서늘한 환경을 선호했을 가능성이 있다. 당시 해발 1000m 미만 지역에서는 오스트랄로피테쿠스 유적이 발견되지 않는다는 점이 이를 뒷받침한다. 이는 침팬지와 유사하게 평균 낮 기온이 25°C이고 밤에는 10°C 또는 5°C까지 떨어지는 지역을 선호했을 수 있음을 의미한다.[38] 현재 말라파 동굴의 고도는 해발 1442m이다.[3] ''A. 세디바''는 ''파란트로푸스 로부스투스'' 및 ''호모 에르가스터''/''호모 에렉투스''와 같은 시기에 살았다. ''오스트랄로피테쿠스 아프리카누스''가 이 시기 즈음 멸종했음을 고려할 때, 남아프리카는 약 200만 년 전 주요 기후 변동과 변동성이 시작되고 ''호모'' 및 ''파란트로푸스''와의 경쟁이 심화되기 전까지 ''오스트랄로피테쿠스'' 속의 피난처 역할을 했을 가능성이 있다.[7]
참조
[1]
웹사이트
"''Australopithecus sediba'' fossil named by 17-year-old Johannesburg student"
http://www.origins.o[...]
Origins Centre
2010-06-04
[2]
논문
"''Australopithecus sediba'' at 1.977 Ma and Implications for the Origins of the Genus ''Homo''"
[3]
논문
"Geological setting and age of ''Australopithecus sediba'' from Southern Africa"
http://doc.rero.ch/r[...]
[4]
논문
"''Australopithecus sediba'': a new species of ''Homo''-like australopith from South Africa"
[5]
논문
"Dental Morphology and the Phylogenetic \"Place\" of ''Australopithecus sediba''"
[6]
논문
"Early ''Homo'' at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia"
[7]
논문
"Contemporaneity of ''Australopithecus'', ''Paranthropus'', and early ''Homo erectus'' in South Africa"
[8]
논문
Candidate human ancestor from South Africa sparks praise and debate
http://doc.rero.ch/r[...]
[9]
논문
Claim over 'human ancestor' sparks furore
http://www.nature.co[...]
Nature News
2010-04-08
[10]
논문
Temporal evidence shows Australopithecus sediba is unlikely to be the ancestor of Homo
[11]
뉴스
Palaeoanthropology: Malapa and the genus ''Homo''
http://www.nature.co[...]
Nature (journal)
2011-10-05
[12]
논문
Hominin Taxonomy and Phylogeny: What's In A Name?
https://www.nature.c[...]
[13]
논문
The Endocast of MH1, ''Australopithecus sediba''
[14]
논문
"''Australopithecus sediba'' and the emergence of ''Homo'': Questionable evidence from the cranium of the juvenile holotype MH 1"
[15]
논문
Mandibular ramus shape of ''Australopithecus sediba'' suggests a single variable species
[16]
논문
Body Size and Proportions of ''Australopithecus sediba''
http://www.paleoanth[...]
[17]
논문
The postcranial skeletal maturation of ''Australopithecus sediba''
https://dspace.lboro[...]
[18]
논문
The cervical spine of ''Australopithecus sediba''
[19]
논문
The Vertebral Column of ''Australopithecus sediba''
[20]
논문
Mosaic Morphology in the Thorax of ''Australopithecus sediba''
[21]
논문
A Partial Pelvis of ''Australopithecus sediba''
[22]
논문
The Upper Limb of ''Australopithecus sediba''
[23]
논문
Adaptation to suspensory locomotion in ''Australopithecus sediba''
[24]
논문
"''Australopithecus sediba'' hand demonstrates mosaic evolution of locomotor and manipulative abilities"
[25]
논문
Rearfoot posture of Australopithecus sediba and the evolution of the hominin longitudinal arch
[26]
논문
The foot and ankle of ''Australopithecus sediba''
[27]
논문
The subtalar joint complex of ''Australopithecus sediba''
[28]
논문
The diet of ''Australopithecus sediba''
[29]
논문
Mechanical evidence that ''Australopithecus sediba'' was limited in its ability to eat hard foods
[30]
논문
Comparative biomechanics of Australopithecus sediba mandibles
[31]
논문
The Lower Limb and Mechanics of Walking in ''Australopithecus sediba''
[32]
논문
Reconstructing birth in ''Australopithecus sediba''
[33]
논문
Distinct growth of the nasomaxillary complex in ''Au. sediba''
[34]
논문
Osteogenic tumour in ''Australopithecus sediba'': Earliest hominin evidence for neoplastic disease
[35]
논문
Evidence of fatal skeletal injuries on Malapa Hominins 1 and 2
[36]
논문
Carnivoran Remains from the Malapa Hominin Site, South Africa
[37]
논문
Botanical remains from a coprolite from the Pleistocene hominin site of Malapa, Sterkfontein Valley, South Africa
[38]
논문
Bipedality and hair loss in human evolution revisited: The impact of altitude and activity scheduling
[39]
문서
남아프리카
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com